E R G E B N I S S E
Allgemeines
Daphniidae produzieren unter populationsspezifischen Umständen,
auf die hier nicht näher eingegangen werden kann (Licht, Temperatur,
Populationsdichte, Stoffwechselendprodukte spielen dabei wahrscheinlich
die Hauptrollen; Stross, 1969; Ferrari et al., 1982; Carvalho et al.,
1983), einen zum Teil verdickten Carapax, in dem bei der nächsten
Häutung ein oder zwei Dauereier zu liegen kommen. Der Carapax
wird dabei nicht, wie allgemein angenommen, sukzessive umgebaut, sondern
schon bei der Vorbereitung der Häutung als Ephippialcarapax angelegt.
Nicht selten kann man unter dem noch nicht gehäuteten alten Carapax,
schon die Anlage des neuen erkennen, der bereits eindeutig die Ephippialskulptur
aufweist (Bild 1).
Aus ökologischen Gründen sind die meisten Ephippien der
Daphniidae schwimmend. Nach der gängigen Meinung (Wolff, 1904;
Flössner, 1972, etc.) wird die Schwimmfähigkeit durch Luft-
oder Gas-Einlagerungen erreicht. Eine Überprüfung dieser
Behauptungen wurde allerdings bislang nicht durchgeführt. Tatsächlich
aber lassen sich aus den Ephippien Luft(?)-Bläschen herausquetschen.
Dies gelingt auch bei ganz frisch abgeworfenen Ephippien und solchen,
die z.B. im Fadenalgengewirr hängen geblieben sind und deshalb
nicht an der Oberfläche trieben. Um genaue Aussagen über
die Genese der Luft- bzw. Gas-Bläschen machen zu können
müßten allerdings eine chemische Analyse derselben und
eine Bestimmung des spezifischen Gewichtes der Ephippien durchgeführt
werden. Tatsache ist auch, daß sich in der Cuticula der Daphniidae
durchwegs Kammern befinden, die sich zumindest an der Oberfläche
vollständig mit Luft füllen und häufig einen starken
Auftrieb verursachen. Die Kammern sind deshalb im folgenden als Auftriebskammern
und die Gesamtheit derselben als Auftriebskörper bezeichnet.
 |
Bild 1: Simocephalus exsoinosus; Querschnitt
eines weiblichen Tieres vor der Häutung: Der neue Carapax
ist für eine Ephippium vorgesehen und trägt bereits
die vollen Ephippialmerkmale. |
Auftriebskörper
Die Auftriebskörper sind fast immer gekammert. Die Kammern sind
zylindrisch, immer mehr oder weniger senkrecht zur Ephippialoberfläche
ausgerichtet. Die Grundfläche der Zylinder zeigt stets Tendenz
zum Sechseck (Wolff, 1904). Bei der Gattung Daphnia sind die Auftriebskörper
annähernd auf das ganze Ephippium verteilt. Der dorsale Kiel
besteht aus kompakten Cuticula-Lamellen. Dorsal und ventral nimmt
die Höhe der Zylinder ab. Lediglich in einem kleinen Bereich
um die beiden Eier, den sog. "Eilogen" ist die Höhe
der Zylinder stark bis völlig reduziert (siehe auch Schultz,
1977). Zwischen diesen Eilogen sind die Zylinder jedoch wohl ausgebildet
Die äußere und innere Cuticula sind dünn, die Skulpturierung
ist durch die gasgefüllten Kammern bedingt (Tafel, Fig. 2).
Von mir gefundene Ctenodaphnia-Arten im burgenländischen Seewinnkel
besitzen Ephippien, die sofort nach der Häutung des Carapax untergehen.
Erwartungsgemäß sind die Auftriebskörper reduziert,
aber dennoch meist gut zu erkennen. Am deutlichsten sind die Auftriebskörper
noch bei Ctenodaphnia magna, am wenigsten deutlich bei C. atkinsoni.
Bei C. magna sind teilweise sogar noch die einzelnen Kammern auszumachen.
Allerdings sind zusätzliche Säulchen eingezogen, die hauptsächlich
quer verlaufen. Sie geben den ehemaligen Auftriebskörpern ein
schwammiges Aussehen. Lokalisiert sind die Auftriebskörper bei
Ctenodaphnia im dorsalen und ventralen Bereich, sowie cranial und
caudal, also ringförmig um die Eilogen. Zwischen den beiden Eilogen
sind von den Auftriebskörpern auch keine Reste mehr zu erkennen.
Die Cuticula ist bei dieser Untergattung sehr dick, auch um die Auftriebskörper
(Tafel I, Fig. 1).
Simocephalus besitzt eine sehr derbe Ephippialcuticula. Auftriebskörper
sind ausschließlich ventral ausgebildet. Im Bereich des Eies,
aber auch dorsal davon sind sie völlig reduziert. Dorso-median
ist eine unpaare, ungekammerte, durchgehende Aufwölbung zu finden
(Tafel I, Fig. 4), in der häufig noch unregelmäßige,
häutige Bildungen, die möglicherweise die Reste eines Kieles
(wie bei der Gattung Daphnia) darstellen. Bei Ceriodaphnia sind die
Verhältnisse ähnlich. Auch hier sind die Auftriebskörper
im Bereich der Eilogen und dorsal des Eies völlig reduziert.
Caudal und teilweise auch cranial des Eies ziehen die Auftriebskörper
bis zur Dorsalkante. Die Kammern der ventralen Auftriebskörper
sind geräumig und eher regelmäßig (Tafel I, Fig. 3).
Die Cuticula ist im gesamten wesentlich zarter, als bei Simocephalus.
Eine krasse Ausnahme bildet Ceriodaphnia megops. Hier sind dorso-lateral
des Eies mächtige, paarige Aufwölbungen vorhanden, die sich
auch von außen gesehen deutlich von der Oberfläche abheben
und Auftriebskörper enthalten. Die gasgefüllten Kammern
sind hier eher schmal und hoch. Die ventralen Auftriebskörper
scheinen gegenüber den anderen Ceriodaphnien etwas reduziert,
die Kammern sind niedriger (Tafel I, Fig. 5).
Dorsale Auftriebskörper finden sich auch durchgehend bei der
Unterfamilie der Scapholeberinae. Bei Megafenestra aurita sind diese
von außen kaum, am Querschnitt aber deutlich zu erkennen. Sie
heben sich kaum von der Ephippialoberfläche ab, führen eher
ringförmig um das Ei, vereinigen sich mit dem ventralen Auftriebskörper
(ähnlich Ctenodaphnia). Die dorsalen Auftriebskörper sind,
ebenso wie die ventralen, mäßig hoch gekammert. Die Cuticula
ist im Bereich der Eilogen sehr dick (Tafel I, Fig. 7). Bei der Gattung
Scapholeberis innerhalb der Scapholeberinae sind die Verhältnisse
etwas anders. Prinzipiell sind auch hier dorsale Aufwölbungen
vorhanden, sie prägen auch das äußere Erscheinungsbild,
wenngleich nicht in so einem großen Maße, wie bei Ceriodaphnia
megops. Die Aufwölbungen sind aber hier, wie sich an den Querschnitten
zeigte, ungekammert. Die ventralen Auftriebskörper sind in der
üblichen Art und Weise ausgebildet und auch deutlich gekammert(
Tafel I, Fig. 6).
TAFEL 1 |
|||
 |
 |
 |
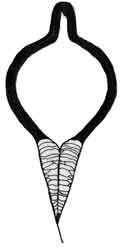 |
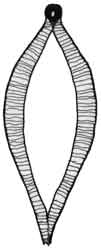
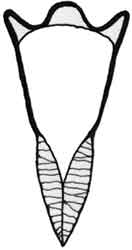
Fig. 1: Ctenodaphnia sp. |
Fig. 2: Daphnia sp. |
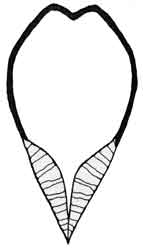
Fig. 3: Ceriodaphnia sp. |
Fig. 4: Simocephalus sp. |
 |
 |
 |
|
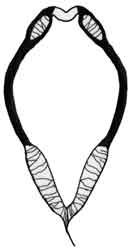
Fig. 5: Ceriodaphnia megops |
Fig. 6: Scapholeberis sp. |
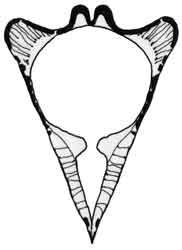
Fig. 7: Megafenestra aurita |
|
Verschlußmechanismus
Nach dem Freiwerden des Ephippiums und Abreißen des restlichen
Carapax an der präformierten Bruchstelle legen sich die beiden
ventralen Ephippialhälften über eine kürzere oder längere
Strecke eng aneinander. Der Druck, der die beiden Ephippialhälften
aneinanderpreßt, muß dabei von der Ephippialhülle
selbst kommen, das heißt, daß sich das Ephippium, bedingt
durch den statischen Aufbau, von selbst verschließt und auch
dadurch geschlossen bleibt. Durch (bisher noch weitgehend ungeklärte)
Quellungsvorgänge wird dieser statische Aufbau dann zerstört
und das Ephippium öffnet sich, um die Jungdaphnie zu entlassen.
An den inneren Flächen der dabei aneinanderstoßenden Hälften
können lediglich, wie bei Ctenodaphnia magna, fädig-fransige
Bildungen vorkommen. Häkchen oder ähnliche Strukturen, die
die Hälften aktiv zusammenhalten, sind allerdings nie zu beobachten.